
water alkalinity
Type of resources
Available actions
Topics
Keywords
Contact for the resource
Provided by
Years
Formats
Representation types
-

Cabras Lagoon is located on the west coast of Sardinia (Italy), in the Gulf of Oristano (39°56’37’’N, 08°28’43’’E), and occupies about 2280 ha, with a mean water depth and maximum of 1.6 and 3 m respectively. The watershed of the site extends over approximately 430 km2. The input of freshwater into the lagoon is scarce and irregular because of the semi-arid Mediterranean climate. Most of the freshwater comes from the small Mare ‘e Foghe River, located in the north. The predominance of agriculture in the region and the release of poorly depurated urban waste account for the high nutrient loads deposited in Cabras Lagoon. The resident population of about 38,000 inhabitants is grouped in 19 urban centres, the largest being Cabras, which is located on the southeast coastal side of the lagoon. During the twentieth century, the lagoon and its watershed underwent several modifications as a consequence of human activities that affected the hydrology and hydraulics of the region. In addition, in the late 1970s, water exchange with the sea was altered by the dredging of a large canal, the Scolmatore (spillway), which connected the lagoon with the adjacent Gulf of Oristano. The canal was constructed to avoid flooding of adjacent land during the heavy rainfall that occurs in winter. In addition, a cement dam was built into the Scolmatore to prevent further increases in the lagoon’s salinity and artificial barriers were constructed to control the fish catch, thereby impeding direct communication between the lagoon and the sea. Now the only link to the sea is via four very narrow creeks that flow into the large canal from the southern part of the lagoon over the barrier. The lagoon has a high economic rating due to ex¬tensive fishery activities, involving about 300 people and those involved in related enterprises. In the site, in addition to the guard houses and warehouses of fishermen, there is also a restaurant, where the products of fishery in the lagoon are offered. In 1998, fish productivity reached 40,000 kg km-2, corresponding to a catch of 850 tonnes but these values fell to around 20,000 kg km-2 and less than 80 tonnes after 1999. In fact, its high trophic status has often exposed the lagoon to important dystrophic crises, which have caused large reductions in its fishing productivity. Scientific monitoring has been carried out since the strong dystrophic crisis that affected Cabras Lagoon during the summer of 1999, killing the whole aquatic biota. A long-term series of data is available and derives from high-frequency measurements and samplings to assess environmental and biological parameters. In particular the data concern the main trophic descriptors (Secchi depth, temperature, pH, conductivity, dissolved oxygen and saturation, alkalinity, NH4-N, NO2-N, NO3-N, total nitrogen, soluble reactive phosphorus, total phosphorus, dissolved silica) and phytoplankton abundances, as chlorophyll a, cell densities and biomass, class and species composition. The activity was interrupted in 2009.
-

Lake Temo (IT10-006-A) is located in north-west Sardinia. The construction of the dam began in 1971 and ended in 1984. The reservoir lies at 226 m a.s.l. and has an area of 4.81 km2, a mean depth of 15.8 m and a maximum capacity of 91 x 106 m3. Its catchment extends for 142 km2.
-

Lago Alchichica se localiza a una elevación de 2300 msnm con una profundidad 62 m y es de agua salobre.Habitan organismos endémicos y se encuentran rocas calizas de particular interés por los microorganismos que viven en ellas. Se encuentra en la Cuenca Oriental de Puebla.
-

Lake Cuga (IT10-003-A) is located in the north-western part of Sardinia. The reservoir was built in 1965, but its first filling was in 1975. It lies at an altitude of 114 m a.s.l. and is extended for about 58 x 106 m2, with a maximum and average depth of 45 m and 11 m, respectively. It has a volume of 34 x 106 m3. The waters are used mainly for irrigation and drinking supplies. Cuga Lake is classified as eutrophic since the early years of its filling.
-

Lake Monte Lerno (IT10-004-A) is located in the North East part of Sardinia in the municipality of Pattada. Its construction was completed in 1980. The catchment area is extended for 160 km2. The reservoir lies at 563 m a.s.l. and has a maximum area of 4.4 km2, a mean depth of 14.9 m and a volume of 89.5 x 106 m3 of water. Its waters are used for drinking and irrigation. Lake Monte Lerno is classified as eutrophic.
-
Runoff and runoff chemistry at LTER Zöbelboden, Austria
-
HYBAM observatory runs a network of 17 stations for hydrological, sediment and geochemistry sampling in the Amazon basin. The infrastructure is shared between the partners in each country including France, Brazil, Peru, Bolivia, Colombia and Ecuador. ADCPs are used for river water flow measurements. Samples are processed in different laboratories at Manaus and Brasilia University (Brazil), Lima University (Peru) and Toulouse University (GET laboratory in France). For comparisons with other large tropical rivers, one stations is maintained along the Orinoco River (Venezuela) and Congo River (Congo) HYBAM data are used in a large variety of programs in relation to hydrology, geomorphology, geochemistry, climatology, climate change, carbon cycle, remote sensing and isotopic tracers. The data produced by the HYBAM observatory are made available online on its website (www.ore-hybam.org) to provide to the research community long term assessments on the hydrology of world largest basin.
-

Lake Cedrino (IT10-002-A) is the result of the dam on the Cedrino River built in 1984. The lake is located in the eastern centre of Sardinia. It has a surface area of 1.5 km2 and a volume of 20 x 106 m3 when it is filled to the maximum share (103 m a. s.l.), and a mean depth of 26.5 m. The catchment covers about 627 km2.
-

The site of the Boknis Eck Time Series (BE) is located at the entrance of the Eckernförde Bay (54°31.2' N, 10°02.5' E) in the southwestern Baltic Sea. It has a water depth of 28 m with muddy sediments. Riverine inputs into the Eckernförde Bay are negligible and thus the overall hydrographic setting at BE is dominated by the regular inflow of North Sea water through the Kattegat and the Great Belt. Seasonal stratification occurs usually from mid-March until mid-September and causes pronounced hypoxia which sporadically become anoxic.
-
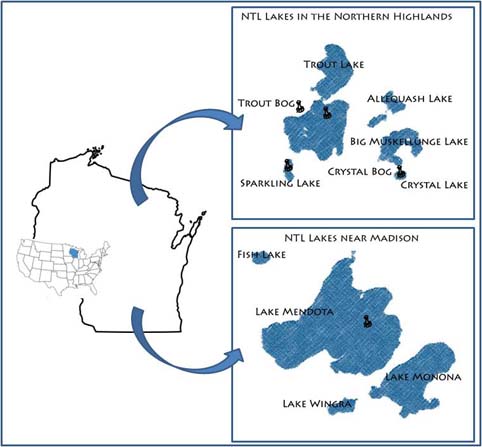
Our primary study sites include a set of seven northern Wisconsin and four southern Wisconsin lakes and their surrounding landscapes. The project, which started in 1981, is administered by the Center for Limnology at the University of Wisconsin-Madison.